|
Neuroscience of Emotion, reward,
pleasure, motivation, decision-making, taste, olfaction, touch, and appetite
including
implications for the control of food intake and obesity
Key
discoveries (see link above) include
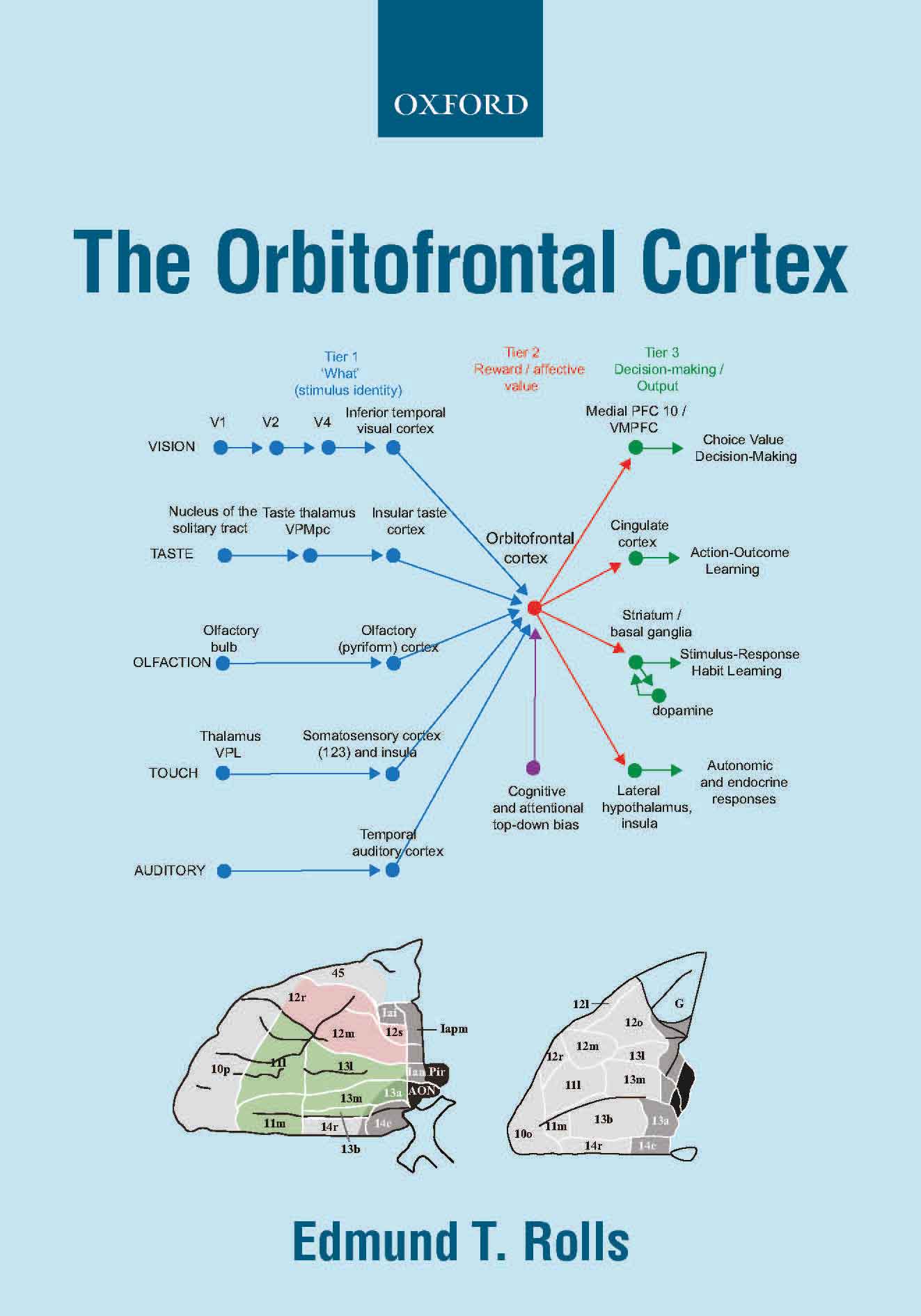
reward value neurons, and non-reward (negative reward prediction error)
neurons, in the orbitofrontal
cortex, leading to a theory of the functions of the orbitofrontal
cortex in emotion, motivation, and depression supported by human
neuroimaging and
neuropsychological investigations; face identity and face expression
neurons in
the orbitofrontal cortex important in social behaviour; the secondary
taste cortex in the
orbitofrontal cortex which represents taste reward value in contrast to
the primary taste cortex in the insula which represents taste identity;
the representation of fat in food by neuronal activity that encodes the
coefficient of sliding friction; olfactory neurons in the orbitofrontal
cortex that represent reward value; food reward value neurons;
reward-specific / sensory-specific satiety which was discovered by
neuronal recordings
from food reward value neurons and is important in food intake control,
but which applies to all reward and no
punishment systems; the orbitofrontal cortex as a key brain region for
brain-stimulation reward; the synaptic basis for reward-specific
satiety and reward-specific or incentive motivation; many discoveries
with functional neuroimaging
and neuropsychology of how these discoveries help to reveal the
functions of the orbitofrontal cortex, anterior cingulate cortex, and
amygdala in reward processing and emotion in humans; the functions
revealed by functional neuroimaging of the human orbitofrontal cortex
in the reward value of the sight, smell, taste and oral texture of
food, and thereby in human appetite control; a focus for reward value
decision-making
in the ventromedial prefrontal cortex revealed by fMRI in combination
with an integrate-and-fire attractor network decision-making model; the
connectivity of the human orbitofrontal cortex and anterior cingulate
cortex in humans including to the hippocampal episodic memory system
for memory and to the dopamine system; and a theory of
emotion, and of motivation, and of their relations to cortical systems
for reasoning.
Neuroscience of Vision
Key discoveries (see link above) in the inferior temporal
visual cortex include face neurons; face expression neurons; invariance
for
size, position, view, lighting, and spatial frequency of face and
object neurons; how information is encoded by sparse distributed
representations in the brain at the single neuron and population of
neurons levels; that face neurons can learn how to represent new faces
in a few trials; that natural scenes greatly reduce the size of face
and object neurons' receptive fields, facilitating vision in natural
environments; a biologically plausible theory
and model (VisNet) of the learning of invariant face and object
neuronal responses in the ventrolateral visual cortical stream;
face expression and head motion neurons in a visual cortical stream to
the cortex in the superior temporal sulcus involved in social behaviour
and implicated in autism; object-based visual motion neurons in the
cortex in the superior
temporal sulcus and how they are computed; how space in cortical scene
areas is represented by
allocentric spatial view cells in for example the parahippocampal scene
area and
hippocampus; the connectivity in humans of a ventromedial visual
cortical stream
to the medial parahippocampal cortex and hippocampus; the theory that
spatial view cells are learned by a ventral stream
feature combination computation and are important in visual scene
representations for memory and
navigation; a theory of how gain modulation
complemented by slow learning enables coordinate transforms in the
dorsal visual system to world-based (allocentric) coordinates used for
self-motion update of parahippocampal and hippocampal spatial view
cells, useful for navigation; how faces, scenes, body parts and tools
have differently lateralised representations, which are also different
in females compared to males; the connectivity in humans (analysed with
effective connectivity, functional connectivity, and diffusion
tractography with fMRI and magnetoencephalography) of four visual
cortical processing streams, a ventrolateral stream for faces and
objects; a stream to the cortex in the superior temporal sulcus for
social stimuli such as face expresions and movements; a ventromedial
stream to the hippocampus for scenes; and the dorsal stream to the
parietal cortex including the human inferior parietal cortex; and the
use of dynamical graphs to show that the whole ventromedial
cortical scene network is separate from the whole ventrolateral
cortical face and object network by measuring flows within each visual
cortical network.
Neuroscience of Memory, Spatial Function, and Navigation
Key discoveries (see link above) include
primate hippocampal and parahippocampal spatial view cells, which
revolutionize our understanding of spatial representations for memory
and navigation in humans and non-human primates by being so completely
different from the rodent hippocampal place cells discovered by O'Keefe
and the entorhinal cortex grid cells discovered by the Mosers; the
allocentric properties of spatial view cells; the prediction property,
related to imagination, of hippocampal spatial cells which respond when
a part of the scene is looked at even when the scene is not visible,
for example in the dark or when obscured by curtains; episodic
memory-like neuronal activity related to one-trial object-location and
reward-location memory storage and recall tasks; a neuronal network
theory of
hippocampal episodic memory and its recall to neocortex, which is the
only quantitative theory of these processes; a theory of
navigation using spatial view cells responding to landmarks; the
spatial view ('Where'), object ('What') and Reward pathways to the
human hippocampus; the human ventromedial
visual cortical stream with the theory that
spatial view cells are learned by a ventral stream feature combination
computation and implement scene representations for episodic memory and
for navigation; hippocampal whole body motion cells, later found in
rodents and termed 'speed cells'; a memory for long-term familiarity by
neurons in the
perirhinal cortex; a theory of the generation of hippocampal time cells
from entorhinal cortex time ramping cells; the discovery that the
human cholinergic basal forebrain neurons receive from the medial
orbitofrontal cortex and the theory that this connectivity, and reward
inputs to the hippocampus, are important in memory consolidation; and
the connectivity in humans of the hippocampal episodic memory system
with semantic memory regions in the anterior temporal cortex and
inferior parietal cortex, with a theory of how these hippocampal inputs
contribute to semantic memory formation. For
working memory and top-down attention, the discovery of the
connectivity of a dorsal prefrontal cortex system with inferior
parietal cortex regions, which complement the dorsolateral prefrontal
spatial and inferior prefrontal cortex object systems.
Computational neuroscience theories of brain
function and
behaviour
Key
computational neuroscience theories (see link above) are
of the hippocampus in episodic memory and navigation;
of how the hippocampus can contribute to the formation of anterior
temporal lobe and inferior parietal cortex semantic memories; of
the ventral stream visual pathways for invariant face and object
recognition (VisNet); of invariant global and object-based motion recognition in the dorsal visual
system;
of the functions of the orbitofrontal cortex in
reward value, emotion, decision-making, and depression; of how
neocortical pyramidal cells can implement the learning of new
categories, attractor memory, and top-down recall and attention; of the
generation of time cells in the hippocampus; of how alterations in the
stability of cortical attractor networks can account for the symptoms
of schizophrenia, obsessive compulsive disorder, depression, ADHD, autism,
normal aging, and creativity; of the computational utility of diluted
connectivity in attractor, pattern association, and competitive
networks in the cerebral cortex; of the relation between the mind and
the brain utilizing levels of explanation in which causality operates
within but not between levels; and of how information in
encoded by a sparse distributed firing rate representation in the brain
based on my neuronal recordings in many cortical regions. A
computational neuroscience approach to understanding brain function is developed in my books Brain Computations and Principles; and AI (2026, Oxford University Press, Open Access) which updates my earlier books Brain Computations and Connectivity (2023, Oxford University Press, Open Access) and Cerebral Cortex: Principles of Operation
(Rolls, 2016, Oxford University Press, .pdf at www.oxcns.org). This
computational neuroscience approach considers what is computed in each
cortical / brain region, how it is computed, how the brain
regions are connected, and provides a fundamental and unifying approach
to understanding how our brains work in health and disease, with
implications for treatment.
Discoveries
on the brain bases of mental disorders
Key
discoveries (see link above) are on depression, in which we find
reduced
connectivity
and activation of the reward-related medial orbitofrontal cortex in
depression; and increased connectivity and activation of the
non-reward-related lateral orbitofrontal cortex, which supports my
theory of depression, and suggests new treatments for depression, for
conventional antidepressant drugs do not work on the medial
orbitofrontal cortex. Key discoveries on Autism Spectrum Disorder are
that the superior temporal cortex face expression and gesture system
which we discovered and which is so important in social behaviour has
reduced connectivity with the orbitofrontal cortex emotion system. Key
discoveries on schizophrenia are that reduced connectivity in some
cortical systems increases the variability and reduces the stability of
some cortical regions; and that there is reduced forward
compared to backprojection cortical connectivity, which alters the
balance of
connectivity and facilitates internally driven instead of externally
driven brain activity. A key discovery on normal aging is about how
reduced
cholinergic activity and reduced connection strengths between neurons
can impair the operation of autoassociation
networks involved in the storage and recall of memory. We have also
shown how disorders of emotion are present in patients with damage to
the orbitofrontal cortex, and how these disorders can be understood in
terms of the functions of the orbitofrontal cortex. In addition, we
have discovered that functional and structural differences of the
orbitofrontal cortex, anterior cingulate cortex, and hippocampus are
associated with lower cognitive performance and with mental health
symptoms including depression, and are associated with childhood
traumatic events, the family environment, low age of the mother,
prolonged nausea and vomiting in pregnancy, and low sleep duration.
Human Cortical Connectivity
Key
discoveries (see link above) in papers published in 2022-2026 are of a
ventromedial
visual cortical stream for spatial, 'where', information to reach the
human parahippocampal scene area and hippocampal memory and navigation
system where spatial view cells are found, which therefore are likely
to be built by a visual feature combination computational process; of
the
pathways for reward and object inputs in humans to the
hippocampal memory
system; of how the hippocampal episidic memory system connects to
anterior temporal lobe and inferior parietal cortex semantic memory
systems; of the role of the orbitofrontal cortex and pregenual anterior
cingulate cortex in providing inputs to the cholinergic basal forebrain
system and thereby modulating memory consolidation;of the role of the
orbitofrontal cortex in providing inputs to the dopamine system; of the
connectivity
of the human dorsal or supracallosal anterior cingulate cortex with
premotor and reward systems enabling it to play an important role in
action-outcome learning, which is a key function of emotion; of the
connectivity of the 'What' and 'Where' parts of the posterior cingulate
cortex important in memory, and of their lateralisation; of the
connectivity of the different
prefrontal cortex systems for different types of working memory; of the
human auditory cortical pathways; of the human somatosensory
pathways which via the insula reach finally the anterior part of the
greatly developed
human inferior parietal cortex; and of the frontal pole cortex with a
theory of how it is involved in exploit vs explore.
|