
B18. 2026
B17. 2026
B16. 2023

B12. 2016
B14. 2019

B9. 2010

B11. 2014

B8. 2008

B10. 2012

B3. 1998
|
Overview: Rolls has developed with colleagues computational neuroscience theories
that are closely linked to his discoveries made in neuronal recording, functional neuroimaging,
and
patient investigation studies.
An aim of the theories has been to go beyond empirical discoveries
about brain function to provide a biologically based framework
for understanding how the brain performs it computations. This
understanding in turn leads to better understanding of and potential
treatments for many disorders of brain function. Rolls' computational
neuroscience approach has included analyses of how information is
represented in the brain (508), of stochastic dynamics in the brain (B9),
and of how memory, visual face and object recognition, and emotion
systems operate in our brains (B18). Key summary descriptions are in B18, B16 and B12.
Brain Computations and Principles; and AI (B18, Open Access) is an update to B16 and B12 that expands the analysis of brain computations greatly to the human
brain by including much evidence on the connectivity of the human
cerebral cortex, which provides a connectional framework for analysing
the computations performed by different regions of the human cerebral
cortex; by adding 16 Chapters on the principles of operation of the
brain; and by extending the comparisons between computations performed
in the brain and in AI. Some of the discoveries are summarized next.
Principles
of Operation of the Cerebral Cortex (B18, B12, B16, 640, 639, 581).
Brain
Computations: what computations are performed, and how they are
performed, in different brain systems.
Rolls has made key contributions to understanding what computations are
performed, and how they are performed, in primates including humans for
many of the systems described in this book (B18, B16).
A biologically plausible approach to computations in the brain using
local synaptic learning rules is described, and contrasted with deep
learning including AI approaches which appear so far not to be
biologically plausible (B18, B16, 684).
Given that the
brain has diluted asymmetrical connectivity, graded firing rates, and
sparse representations, extensions with Alessandro Treves of the theory
of attractor networks beyond the Hopfield approach, and pattern
association networks, to show that these more realistic brain-like
systems have high capacity, are stable, and operate rapidly within 2.5
time constants of the synapses (B3, 154, 150, 186, 222, 228, 235, 686, 692, B16, B18). These are now fundamental building blocks of current approaches to biologically plausible cortical computations (B16, B18).
A
theory of how neocortical pyramidal cells can implement the learning of
new categories, attractor memory, and top-down recall and attention
(640, B16, B18).
A
theory of emotion, and pleasure, and reward; and the principles of
their
implementation in the brain (B5,
B11,
B13, 273, 520,
148, 364, 428, 509, 526,
533, 534,
552,
579, 674, 697, B16, B18).
A
theory of motivation, and its relation to reward and emotion systems (557, 674, 697, B18).
In this context, a key
principle in primates including
humans is that reward value and emotional valence are represented in
the orbitofrontal cortex (and amygdala), whereas before that, the
representations are about objects and stimuli independently of value,
in the inferior temporal visual cortex, the primary taste cortex in the
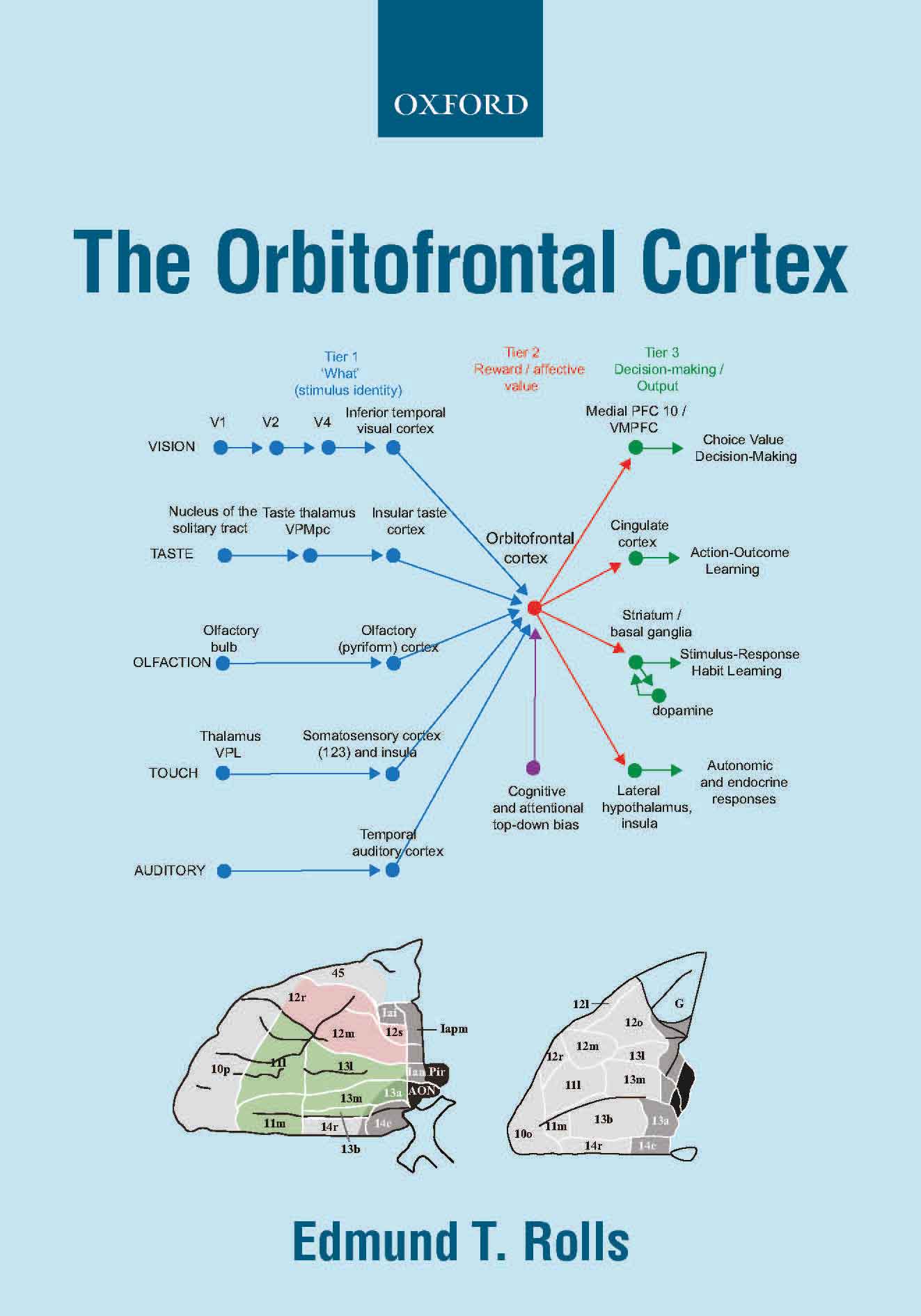
insula, and in the olfactory cortex (B11, B13, B14, B16, 674, B18). This provides for the separation
of emotion from perception in primates including humans.
In this framework, the dopamine
neurons are seen
as receiving their information from brain regions such as the
orbitofrontal cortex, via the ventral striatum and habenula (572,
B11, B13, B14, B16, 649, B18).
Further, orbitofrontal cortex neurons encode reward value and hence
emotion, independently of goal-related actions. The orbitofrontal
cortex provides reward-related information to the cingulate cortex for
action-outcome learning, and to the basal ganglia for habit-based
responses (B11, B13, B14, 579, 606, 674, B16, B18).
The
roles of the emotional and the reasoning systems in decision-making (497, 518, 674, 697,
B5, B10, B11, B13, B16, B18) including in
economic decision-making (B11, 600).
A
biased activation theory of top-down attentional and cognitive control (339, 488, 520, 530, B11,
B12, 603, B16, B18).
A
theory and model of hippocampal operation and episodic memory (111, 125,
136, 163, 186, 200, 205, 258, 266, 268, 300, 306, 307,
309, 345, 370, 403, 411, 415, 433, 453, 479, 521, 527,
529,
539,
545,
550,
584, 643, 672,
B12, B16, 686, 692, B18). This remains
the only quantitative theory of the storage and recall of memories in
the hippocampo-cortical system (686, B18).
A theory for how
hippocampal spatial view cells are involved in memory and navigation (584, 594, 539,
B12, 612, 633, 662, B16, 686, 692, 702, B18) and for the pathways involved in humans (682, 686, 692, 695, B18). A
key concept is that the 'where' spatial view representations in the
primate including human hippocampus are built by feature combinations,
then attractor networks and gain modulation by gaze in a ventromedial
visual cortical stream to the primate including human hippocampus, very
different from the place
representations in rodents (702, 696, 692, 686, 662, 682, 370, 393, B18). These discoveries of the importance of spatial view representations in
the primate and human hippocampus, and the pathways involved, are
revolutionizing (702) our understanding of the functions of the medial
parahippocampal cortex and hippocampus in primates and humans for
memory and navigation in which viewed locations and landmarks in scenes are important (696, 692, 662, 682, 633, 672, 594, 539, 686, B18).
In contrast, given the representations of the place where
the individual is located that are found in the rodent hippocampal and
medial entorhinal cortex system, navigation in rodents is considered to
involve blind path integration from place to place using head
direction, the speed of travel, and the time (662, 682, 672, 686, 702, B18).
In addition, I have shown how the hippocampal
episodic memory system has connectivity to anterior temporal lobe semantic
multimodal including visual regions, and to semantic inferior parietal visual
cortical regions, and have produced a theory and model of how the
hippocampal episodic memory inputs inputs could help to form
semantic memories (694, B18).
A theory and model of the
generation of time in the hippocampal memory system. Entorhinal cortex
time ramping cells produce through a competitive network hippocampal
time cells, providing neuronal mechanisms to encode the order of events
(605, B16, B18).
The theory shows how cells could be generated that show 'replay' and
'reverse replay' (605).
A theory and model of
coordinate transforms in the dorsal visual system using a combination
of gain modulation and slow or trace rule competitive learning. The
theory starts with retinal position inputs gain modulated by eye
position to produce a head centred representation, followed by gain
modulation by head direction, followed by gain modulation by place, to
produce an allocentric representation in spatial view coordinates
useful for the idiothetic update of hippocampal spatial view cells (612). This is useful in navigation using spatial view cells when the view details are obscured (B15, 633, 662, B16, 686, 691, B18).
A dynamical systems approach using
integrate-and-fire neurons to neocortical-hippocampal-neocortical
function that shows that memory recall to the neocortex takes in the
order of 100 ms, which is sufficiently rapid for the backprojections to
be modified using storage in order to implement later recall to the
neocortex (689). The operation of the whole network has implications
for how the hippocampus is involved in the formation of new neocortical
semantic memories.
A
theory of how spatial view cells and hippocampal attractor networks are
involved in the art of memory (using the method of loci) (571,
595).
A
theory and model, VisNet, of invariant visual object recognition in the ventral
visual
system (162,
179,
192,
226,
245,
275,
277,
280, 283, 290, 304, 312, 396, 406, 414, 446, 455, 473,
485, 516, 535,
536,
554,
589,
639, B12, B16, 703, B18).
This approach is unsupervised, uses slow learning to capture
invariances using the statistics of the natural environment, uses only
local synaptic learning rules, and is therefore biologically
plausible in contrast to deep learning approaches with which it is
contrasted (639, B16, 703, B18).
A further advance towards biological plausibility is use of a synaptic
learning rule with, in addition to long-term potentiation,
heteroynaptic long-term depression that depends on the strength of the
synaptic weight, which removes the need for normalization of synaptic
weight vectors, and improves performance (703, B18).
A
theory of invariant global and object-based motion recognition in the dorsal visual
system
(413, B16, B18).
A
theory of the utility of the stochastic dynamics of attractor networks in the
brain for
decision-making and many other aspects of our behaviour, including
creativity
(404, 463, 477, 483, 491, 500, 502, 504, 507, 513, 518, 540,
B9,
B11, B12, B16, B18).
The
stochastic dynamics arises from the almost random
(Poisson) firing times of cortical neurons for a given mean firing
rate, and
gives rise to the concept of The
Noisy Brain (B9, B18) .
Theories
of how alterations in the stability of cortical attractor networks can
account
for the symptoms of schizophrenia (431, 436, 450,
490, 503, 629, 631), of
obsessive compulsive disorder (449), of depression (559,
572, 626, 679), of ADHD (629), of normal aging (540, 613),
and for creativity (585).
The
theories (B12, B13, B14, B16, 681, B18) have implications for
treatment, and are complemented by
neuroimaging investigations (538,
541,
563,
564,
565,
583, 585,
587,
588,
590,
591,
592,
596, 602,
615, 626, 629, B14, B16, 679, 705, B18).
A theory of avalanches in the brain and the temporal variability of functional connectivity (630, 629).
A
non-reward attractor theory of depression (559,
572, B13),
supported by altered connectivity and activation of the orbitofrontal
cortex in depression (564,
583,
588,
590, 591,
592,
596, 602,
615,
616,
623, 626, B13, B14, B16, 679, B18), and a model of non-reward computation in the
orbitofrontal cortex (562).
The
roles of cortical attractor networks in short-term memory and top-down
attention
(294, 295, B6, 47, 360, 372, 379, 391, 410, 520, B8, 523,
530, B12, 640, B16, B18).
The
design of neural networks in the cortex by genetic evolution (284, B12, B16, B18).
Separate limbic systems for emotion and memory, but no single limbic
system (531).
A conceptual framework for understanding the cingulate cortex, how
it forms a part of different limbic systems (606, 649, 661, B16, 674, B18) and the functions of the anterior cingulate cortex (649, 674) and posterior cingulate cortex (661).
A theory supported by effective connectivity findings in humans of
how the orbitofrontal, ventromedial prefrontal, and anterior cingulate
cortex provide a reward value input to the hippocampal memory system,
and by also influencing basal forebrain and septal cholinergic neurons
facilitate memory and if damaged contribute to memory impairments
produced by vmPFC damage (649, B18).
The
representation of information in the brain using a neuronal firing rate code (172, 196, 204, 225, 227, 321,
255,
419,
474, 508, 231, 265, 329, 348, 351, 369, 517, 193, 197, 257, 407, 553,
561,
B12, B16, B18).
The computational utility of diluted
connectivity in attractor, pattern association, and competitive
networks in the cerebral cortex (504,
515,
545,
550,
B12, B16, B18).
A possible implementation of
syntax in the brain using linked attractor networks (537, B16, B18).
This is complemented by the analysis of effective connectivity in the
cerebral cortex, which identifies a 'what and reward' semantic system
involving ventral superior temporal sulcus cortex, the temporal pole,
inferior parietal PGi and orbitofrontal cortex; and a visual face and
object motion and auditory semantic system in the dorsal superior
temporal temporal sulcus cortex especially implicated in social
semantics (654).
Both semantic systems have effective connectivity to Broca's area 44
and 45, which in humans has links to other nearby inferior frontal
cortex regions that are proposed to provide attractor networks for
syntactic computations (654).
A higher order syntactic thought (HOST)
approach
to consciousness (239,
341, 355,
398, 422, 432, 456,
493, 497, 525, 618,
B11, B12,
B13, B16, B18).
A theory of the relation between the mind and the brain utilizing
levels of explanation in which causality operates within but not
between levels (618, 632, 637, B16, B18).
Biological underpinnings of
art and aesthetics (B10, 492, 509,
532,
556,
574).
|